
Der Cortex Cerebri = Die Großhirnrinde
Copyright © by V. Miszalok, last update: 2009-06-14

Let me know
what you think
| Home | Index of Lectures | PDF Version of this Page |
|
Der Cortex Cerebri = Die GroßhirnrindeCopyright © by V. Miszalok, last update: 2009-06-14 |
Let me know what you think |
Die Biologie definiert "Wahrnehmung" sehr extensiv als jedwede Veränderung der Innenwelt, die durch die Außenwelt hervorgerufen wird. Folglich besitzt nicht nur jede Pflanze, sondern jede Zelle Wahrnehmung. Es ist unmöglich den Begriff einzuengen, wenn man willkürliche Begrenzungen vermeiden will (etwa auf Säugetiere oder Menschen). Wahrnehmung gibt es also immer und überall.
Dieser so verwässerte Begriff ist für die Biowissenschaften unbrauchbar geworden und deshalb dort verschwunden, wie es auch anderen Begriffen ergangen ist wie Bewusstsein und Intelligenz und übrigens auch deren Gegenbegriffen Bewusstlosigkeit und Dummheit.
In der Informatik existierten die Begriffe Wahrnehmung und Bewusstsein noch nie. Der Empfang einer 0 oder einer 1 ändert den inneren Zustand eines Computers. Es klingt lächerlich, wenn man sagt, der Computer habe die 0 oder die 1 "wahrgenommen".
Leider hat sich der Begriff Intelligenz in der Informatik erhalten. Hartnäckig hält sich das Gerücht, dass Computer intelligent sein könnten, ja es gibt sogar das Forschungsgebiet "Künstliche Intelligenz". Tatsache aber ist, dass es nur oberflächliche Analogien zwischen Computer-Aktivität und Gehirn-Aktivität gibt. Der Begriff Intelligenz hat in der Informatik eigentlich nur Unheil angerichtet; er hat zu vielen Missverständnissen und zur Verschwendung enormer Summen an Forschungsgeldern geführt.
Bleiben wir bei der Umgangssprache. Die Organe der Wahrnehmung sind offensichtlich die Sinnesorgane: Auge, Ohr, Schmerzrezeptoren etc. Zweifellos gehören auch die Teile des Gehirns dazu, die Signale von diesen Sinnesorganen erhalten. Die Eintrittsplattform für solche Signale ist das sogenannte thalamo-corticale System bestehend aus
1. dem Thalamus, ein paariges 2x kirschengroßes Zellgebiet unten auf der Schädelbasis,
2. der dünnen äußeren Hüllschicht des Gehirns = Neocortex oder kurz Cortex und
3. den reziproken thalamo-corticalen Leitungsbahnen.
Sinnesorgane und Thalamus finden wir im Tierreich fast überall, aber einen Cortex haben die meisten Arten nicht. Säugetiere besitzen einen Cortex. Die Primaten besitzen einen großen Cortex und der Cortex des Menschen ist mindestens dreimal größer als der des ihm am nächsten stehenden Primaten.
Offensichtlich ist dieser große Cortex die materielle Basis der besonderen menschlichen Fähigkeiten wie Abstraktion, Sprache, Intelligenz etc.
Der Cortex bedeckt das gesamte Großhirn in Form einer ca. 2 mm dicken, grauen Haut = Gehirnrinde. Würde man den Cortex vom Gehirn ablösen und ausbreiten, hätte er in etwa die Fläche des Papierformats DIN A3 ≈ 42 x 30 cm.
Grob unterscheidet man sensorische (20%) und motorische (20%) Cortexgebiete und solche die nicht mit der Außenwelt verbunden sind = Assoziationsareale (60%), die weder sensorische noch motorische Aufgaben haben.
| Seitenansicht der linken Hemisphäre links→Stirn, rechts→Hinterkopf Nur ein Teil des (gelben) Sehareals ist sichtbar. Der größere Teil ist in der sagittalen Symmetriefurche versteckt. | 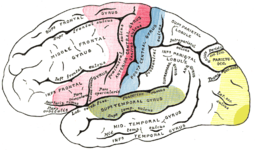 |
rot: motorische Areale blau: sensorische Areale grün: Hören gelb: Sehen weiß: Assoziationsareale |
Der Cortex ist ein Flickenteppich von meist kleinen funktionellen Arealen (maximale Fläche einer 10-Cent-Münze).
Die großen Flächen wie die Sehrinde (gelb ca. 10x10 cm am hinteren Pol) enthalten multiple kleine Areale.
Die Lage der Areale ist bei allen Säugetieren ungefähr gleich, es gibt jedoch eine beträchtliche Artendifferenz: z.B. Raubtiere haben in der Regel einen viel größeren visuellen Cortex als Vegetarier.
Es gibt auch große individuelle Differenzen innerhalb einer Spezies:
Beispiel: Blind geborene Menschen haben keinen visuellen aber einen großen auditorischen Cortex.
Ausdehnung und Lage der Areale sind beim Menschen bemerkenswert flexibel.
Nach Zerstörung eines Areals können kindliche Cortices die fehlende Funktion in der Nachbarschaft neu erlernen (auf Kosten der dort früher residenten Areale).
Die großen Flächen der oben erklärten Areale sind weiter unterteilt in Teilareale und Teilareale von Teilarealen, etwa wie die Bundesrepublik in Länder, Regierungsbezirke, Landkreise und Gemeinden unterteilt ist.
Beispiel: Der visuelle Cortex (=Bundesland) enthält einen primären, einen sekundären und einen tertiären visueller Cortex (=3 Regierungsbezirke). Diese 3 Regierungsbezirke sind zwar ungefähr gleich groß und anatomisch ziemlich identisch, aber sie verarbeiten die visuellen Signale auf 3 verschiedenen Abstraktionsniveaus. Das Abstraktionsniveau steigt vom primären visuellen Cortex (kartographische Codierung der Außenwelt) zum sekundären (symbolische Codierung) zum tertiären (Codierung von Bedeutungen). Der Signalstrom bewegt sich vom primären über den sekundären zum tertiären visuellen Cortex (mit sehr starken Rückkopplungen).
Innerhalb der Regierungsbezirke liegen die Kreise = Teilareale von Teilarealen und in diesen die Gemeinden = Microareale. Auch diese Gemeinden unterscheiden sich in ihren Abstraktionsniveaus. Insgesamt hat jede Gemeinde = Microareal ihren Platz in der militärisch straffen funktionellen Hierarchie innerhalb ihres Teilareals. Diesen Platz in der Hierarchie nummeriert man von 1 bis n.
Zwar kennt niemand die Anzahl n der Stufen im visuellen Cortex noch kann jemand die Verwaltungsgrenzen eines Microareals konkret zeigen, aber wir wissen mit Sicherheit, dass diese Verwaltungseinheiten existieren und dass diese Verwaltungseinheiten nicht gleichwertig sind, sondern dass jede auf ein bestimmtes Niveau einer n-stufigen Signalumwandlungskette spezialisiert ist.
Lassen Sie uns die Kette mit einem Laufindex i nummerieren, wie Informatiker das tun:
So bilden die Microareale Mäander und diese Mäander bedecken insgesamt lückenlos kompakt ein Areal.
Jedes Microareal besitzt mindestens 4 Kabel nach außen. Stellen Sie sich jedes Kabel etwa so vor wie das vieladrige Flachbandkabel, das Ihre Festplatte mit Ihrem Motherboard verbindet.
Kabel 1: Transportiert Signale von i-1 nach i → Aufwärtskabel
Kabel 2: Transportiert Signale von i nach i-1 → Abwärtskabel
Kabel 3: Transportiert Signale von i nach i+1 → Aufwärtskabel
Kabel 4: Transportiert Signale von i+1 nach i → Abwärtskabel
Kabel 5 etc: Weitere Kabel von und zu weit entfernten Zielen.
Unter Signal versteht man den kollektiven elektrische Zustand aller Adern eines Kabels zu einem bestimmten Zeitpunkt t.
Die Bedeutung der Rückkopplung erkennt man daran, dass die beiden Abwärts-Kabel 2 und 4 mehr Adern haben als die beiden Aufwärts-Kabel 1 und 2.
Jedes Microareal besitzt weiterhin ein eigenes Microgedächtnis. Den Inhalt des Microgedächtnisses nennt man Abstraktion oder Prototyp oder Vorurteil.
Je höher ein Areal in der Hierarchie steht, um so verdichteter, pauschaler und abstrakter sind seine Prototypen = Vorurteile.
Aufstieg sensorischer Signale durch die Hierarchie:
Ein Signal wird solange verdichtet und die Hierarchie hinauf geschleust, bis es einem Prototypen = Vorurteil zugeordnet werden kann.
Abstieg durch die Hierarchie:
Nach der Zuordnung produziert der Prototyp ein Signal, das er optimal findet, dieses steigt die Hierarchie hinab und wird darauf untersucht, ob es dem Originalsignal genügend genau entspricht. Falls nicht, wird der Aufstieg wiederholt.
(Looping durch die Hierarchieebenen, bis ein passender Prototyp gefunden wird).
Es ist schwer zu begreifen,
1. warum der Cortex überall gleich dick ist und nicht dort dicker ist, wo viele Abstraktionsniveaus notwendig sind.
2. warum die Abstraktionsniveaus wie Flächenstaaten nebeneinander und nicht übereinander geschichtet liegen.
3. warum die Evolution diesen Weg der flächigen Ausbreitung gewählt hat, statt der Verdickung des Cortex (= vertikale Stapelung der n Abstraktionsniveaus).
Der Nachteil dieser flächigen Architektur liegt auf der Hand: Es ist viel horizontale Verkabelung notwendig mit der fatalen Folge: Die Spezies Mensch kann nicht klüger werden, weil unsere Cortex-Fläche nicht vergrößerbar ist (allenfalls durch weitere Fältelung der Gehirnoberfläche).
Ein Kubikmillimeter Cortex enthält ca. 70 000 Pyramidenzellen unterschiedlichen Typs, 20 000 sonstige Nervenzellen und Interneurone und jede Nervenzelle besitzt ca. 20 000 Synapsen. Weiterhin enthält jeder Kubikmillimeter eine interne Verkabelung von ca. 4 km Länge.
 | Computersimulation aus dem Blue Brain Project des zelluläre Filzes innerhalb einer Cortex-Säule Die Cortex-Zelle hat viele kurze Arme (Dendriten) und einen langen Arm (Axon). Die Arme empfangen oder senden elektrochemische Signale. Die Signale verstärken (exzitatorische Signale, Botenstoff Glutamat) oder schwächen (inhibitorische Signale, Botenstoff GABA) die Erregung fremder Zellen. Bildquelle: Blue Brain Project Gallery |
Unabhängig von der lokalen Funktion und der Hierarchiestufe hat der gesamte Cortex einen erstaunlich einheitlichen laminarer Aufbau aus 6 unterscheidbaren Zellschichten, die traditionell so nummeriert sind: außen S1, innen S6:
Laminierung:
S1 = außen = keine Zellkörper, nur Oberflächen-parallele Fasern
S2 + S3 = dicht gepackte Pyramidenzellen
S4 = Sternzellen
S5 = Durchgangsgebiet mit manchmal extragroßen Pyramidenzellen
S6 = innen = mehrere spezielle Zelltypen
Die Schichten S1 bis S6 sind vertikal stärker vernetzt und arbeiten vertikal intensiver zusammen als horizontal = Säulenarchitektur.
Die Zellen einer Mikrosäule entstehen embryonal durch Zellteilung aus einer Vorläuferzelle. Der Cortex besteht so aus einigen 100 Millionen Mikrosäulen von 2 mm Länge und der Dicke eines Haares.
Benachbarte Mikrosäulen können sich wie bei einem Pinsel zu stärkeren Makrosäulen zusammenschließen = funktionelle Einheiten senkrecht zur Oberfläche, Dicke ca. 0,5 mm. Eine Makrosäule beherbergt ein Teilareal (siehe oben im Kapitel Hierarchien).
Die S1-Schicht bedeckt die Macrosäule = Pinsel = Teilareal horizontal und verbindet Micro- und Makrosäulen (sowohl im Nahbereich wie auch über sehr weite Distanzen).
Folgende Statistik zeigt, wie wichtig die S1-Schicht ist:
ca. 80% aller Synapsen einer Mikrosäule sind Synapsen mit S1-Fasern.
ca. 10% aller Synapsen verbinden vertikal S2 mit S3 mit S4 mit S5 mit S6.
ca. 10% aller Synapsen sind Verbindungen von und zum Thalamus.
Ein Reiz aus der Tiefe des Gehirns aktiviert meist die ganze Säule.
Die Signale von den Sinnesorganen oder von i-1 kommen nicht, wie man erwarten könnte, in S6 an, sondern in S4. Sie steigen von der Ankunftsplattform S4 auf zu S3 und S2 und steigen von oben wieder ab nach S5 und S6.
| S1 | horizontaler Verteiler für Fern-Signale und horizontaler Informationsfluss von Mikrosäulen zu ihren Nachbarn. |
| S2, S3 | Abfahrtsplattform des Aufwärtskabels 3 (i → i+1) und Ankunftsplattform des Abwärtskabels 4 (i+1 → i). |
| S4 | Ankunftsplattform des Aufwärtskabels 1 (i-1 → i). |
| S5 | Bei den meisten Teilarealen verkabelt S5 nur von und nach S1. Wenn das Teilareal mit Motorik zu tun hat, dann ist hier aber die Abfahrtsplattform von Kommandosignalen an die Muskulatur und deren Kopien an den Thalamus. |
| S6 | Abfahrtsplattform des Abwärtskabels 2 (i → i-1). |
Beispiel:
Es gäbe für eine Aufgabe (z.B. Musik hören) n Makrosäulen = Teilareale, wobei die i.te Makrosäule für das Abstraktionsniveau i spezialisiert sei.
Die erste Säule erhält ihren S4-Input roh direkt vom Innenohr.
Die letzte = n.te Säule ist spezialisiert auf das höchste Abstraktionsniveau: Erkennen von Symphonien, Dirigenten, Orchestern, Solisten.
Vertikale Verbindungen innerhalb einer Microsäule:
a) Schichten werden in beiden Richtungen übersprungen und die beiden Informationsflüsse der beiden Richtungen verzahnen sich kammartig.
b) Die Verkabelung ist asymmetrisch. Es ziehen mehr Fasern nach unten (Richtung S1 → S6) als nach oben.
Der Rechenweg rückwärts von abstrakt nach konkret ist offensichtlich schwerer als der vorwärts von konkret nach abstrakt.
Horizontale Verbindungen zwischen den Microsäulen:
Es gibt schwache horizontale Verbindungen in S2 bis S6 von jeder Microsäule zu den unmittelbaren Nachbarn. Diese sind wahrscheinlich durchgehend inhibitorisch und verhindern unkontrollierte flächige Erregungsausbreitung. Die wirklich Signal-tragenden Verbindungen zwischen den Säulen verlaufen alle über S1.
Unsere willkürlichen Bewegungen werden geplant und ihre komplizierten Abläufe werden codiert von den motorischen Arealen des Cortex.
Aber die meisten Bewegungen sind nicht willkürlich und viele davon werden nicht vom motorischen Cortex gesteuert, sondern vom sensorischen Cortex und von den Assoziationsarelen.
Beispiel: Der visuelle Cortex dekodiert nicht nur die beiden Netzhautbilder, sondern erzeugt auch die Augenbewegungen.
Man darf sich von den Begriffen "motorischer" und "sensorischer" Cortex nicht irreführen lassen. In Wirklichkeit existiert keine so klare Trennung zwischen Sensorik und Motorik, wie die Begriffe suggerieren. Beide Aktivitäten sind jedenfalls beim Sehvorgang eng verzahnt.
Der visuelle Cortex sieht etwas, generiert daraus eine Augenbewegung und eine Prognose über das nächste Bild. Dann vergleicht er dieses Bild mit der Prognose und errechnet aus den Differenzen einen neues Sprungziel für die Augenachsen usw.
Die Außenwelt wird also abgetastet durch eine schnelle Abfolge von:
1) Bildaufnahme →
2) Bewertung der Differenz zwischen neuem Bild und alter Prognose →
3) neues Sprungziel (=Sakkade) und neue Prognose.
4) weiter bei 1).
Visuelle Sensorik und visuelle Motorik sind untrennbar ein einziger Vorgang und unser gesamtes visuelles Gedächtnis speichert wahrscheinlich keine Bilder, sondern Folgen von Augenbewegungen.
Der Cortex sendet seine motorischen Befehle an die externe Muskulatur aber 1:1-Kopien dieser Befehle gehen parallel an eine zweite Gehirn-interne Adresse: Den Thalamus.
Der Thalamus ist ein paariges, gut abgrenzbares Gebiet, das je kirschengroß beidseits der Symmetrieebene auf der Schädelbasis liegt. Über seine Funktion ist so viel bekannt:
1) Er erhält Kopien aller aus dem Cortex stammenden Steuersignale zur Steuerung der willkürlichen Muskulatur.
2) Der Thalamus sendet zeitversetzt diese motorischen Signale fast unverändert zurück an Schicht S1 des Cortex.
3) Der Cortex erhält somit seinen eigenen Output über den Thalamus zeitversetzt zurück.
4) Diese verzögerte Rückkopplung dient dem Cortex dazu, den Erfolg seiner motorischen Kommandos zu kontrollieren.
5) Die Erfolgskontrolle misst die Differenzen zwischen dem Plan und der unmittelbar darauf folgenden Ausführung.
6) Ungewöhnliche Differenzen werden im Thalamus emotional bewertet und lösen Alarm aus.
7) Dieser Alarm ist eine notwendige Bedingung für das, was wir Bewusstsein nennen. Thalamus-Schädigung führt zum apallischen Syndrom = Wachkoma.
8) Neben der Verbindung der Cortexareale über die Faserschicht S1 ist die Rückkopplung über den Thalamus ein wichtiger Kommunikationsweg weit auseinander liegender Cortexareale.
9) Die Fasern vom Thalamus durchqueren große Teile der Schicht S1 und informieren "die Linke, was die Rechte gerade tut".
10) Wegen der hohen Informationsflüsse spricht man vom "thalamo-corticalen System" und nennt die reziproken (hin- und zurück) Verbindungen die "cortico-corticalen Bahnen".
Neben dem thalamo-corticalen System gibt es zwei weitere Großstrukturen im Gehirn, die den Cortex beeinflussen:
1) Die "polysynaptischen Schleifen" die, ausgehend von den Basalganglien, über den Thalamus und den Cortex wieder zurück zu den Basalganglien ziehen. Diese Schleifen enthalten vorwiegend inhibitorische GABA und Dopamin ausschüttende Neurone. Wegen der Disinhibition = Schwächung von Schwächung können die Schleifen auch erregend wirken. Sie steuern die nicht-bewusste Motorik wie z.B. Fahrradfahren = prozedurales Gedächtnis und sind beteiligt an der Bildung von Wahrnehmungskategorien.
2) Die "Bewertungssysteme" sind kleine Kerne, deren Axone nicht an Zielzellen andocken, sondern ungezielt weit entfernt in den Extrazellulärraum erregende oder hemmende Substanzen ausschütten (Modulation). Die euphorisierenden Drogen wirken über diese Bewertungssysteme.
Beispiele:
- Locus coeruleus im Gehirnstamm → Noradrenalin
- Raphe-Kerne → Serotonin
- cholinerge Kerne → Acetylcholin
- dopaminerge Kerne → Dopamin
- Hypothalamus → Histamin
- Gehirnstamm → Neuropeptid Enkephalin
- Amygdala moduliert u.a. Hypothalamus, Locus coeruleus
Ca. 50% der Gehirnmasse besteht aus Leitungsbahnen unterhalb des Cortex = Kabelbäume = weiße Substanz. |
 |
Die frappante Tatsache, dass der Cortex uniform und homogen überall aus gleichen Zelltypen 6-schichtig aufgebaut ist und dass man mikroskopisch nicht unterscheiden kann, ob ein Stück Cortex für Sehen oder Hören oder für sonst etwas spezialisiert ist, macht plausibel, dass ein einheitlicher Algorithmus überall im Cortex ausgeführt wird.
Theorie von Vernon Mountcastle 1978:
Die Rechenprozesse sind identisch auf der gesamten Cortexfläche in allen funktionellen Arealen. Es gibt nur einen corticalen Algorithmus, der parallel überall ausgeführt wird.
Was ist ein Prototyp ?
Ein Prototyp ist ein Micro-Gedächtnisinhalt eines Teilareals darüber, wie ein gutes Signal aussehen würde, wenn es vollständig und ungestört wäre.
Jedes Teilareal i vergleicht das via Aufwärtskabel 1 in S4 ankommende Signal mit seinem Prototypen und stellt den Grad der Ähnlichkeit fest.
Ist die Ähnlichkeit hinreichend groß, erklärt sich das Teilareal für zuständig und teilt dies seinem Vorgänger i-1 und seinem Nachfolger i+1 und anderen Nachbarn mit. Andernfalls erklärt es sich für unzuständig und reicht das Signal unverändert nach i+1 weiter.
Beispiel: Das Wort "Gatenschluch" ist unverständlich ohne das Vorhandensein eines Prototypen "Gartenschlauch".
Prototypen sind die Vorurteile, die uns die Evolution kollektiv eingeprägt hat oder die wir individuell durch Erfahrung konstruieren. Schon embryonal besitzen wir ein enormes Vorwissen über die Welt. Diesen Vorrat an Vorurteilen ergänzen wir individuell im Laufe unseres Lebens. Wahrnehmung bedeutet das Wiederfinden von Vorurteilen im chaotischen Datenstrom, den die Sinnesorgane liefern. Wir sehen nur die Dinge, von denen wir Prototypen besitzen. Alles andere verschwindet spurlos.
Vorurteile sind lebensnotwendig für Systeme, die gestörte hochredundante Signale verarbeiten. Solche Systeme müssen Ähnlichkeiten erkennen und bewerten und Voraussagen darüber machen, was wahrscheinlich als nächstes Signal ankommt. Siehe: Redundancy.htm
In diesem Sinne sind Vorurteile nichts, was man verurteilen sollte. Sie sind ein hohes Gut in einer Welt voller Redundanz, Störung und Zufall.
Wir verdanken sie der Evolution oder wir müssen sie durch Versuch und Irrtum mühsam erwerben. Der Umfang unserer Sprache entspricht wahrscheinlich unserem Vorrat an Prototypen. Ein Gehirn ohne Vorurteile wäre sprachlos und leer.
Gleichzeitigkeit
Viele Microareale arbeiten am aktuellen Datenstrom gleichzeitig mit hoher zeitlicher Präzision. Erkennt ein Microareal seinen Prototypen, dann wird es rhythmisch aktiv. Erkennen mehrere benachbarte Microareale gleichzeitig ihre Prototypen, verstärken sie sich gegenseitig. Sie beginnen zu oszillieren, kommen in Resonanz und schwingen synchron. Es entsteht innerhalb von ca. 150 Millisekunden ein synchron taktendes Aktivitätszentrum von etwa tausend Neuronen. Diese synchrone Aktivität signalisiert den nachgeschalteten Neuronen Konsens darüber, dass diverse Signale der Außenwelt zusammengehören.
Wenn ein weiteres entferntes Aktivitätszentrum gleichzeitig schwingt, versuchen sich diese beiden Zentren z.T. über große Entfernungen hinweg zu synchronisieren, d.h. in Gleichtakt zu bringen. Gelingt dies, so entsteht ein Funktions-Zentrum höherer Ordnung. Oszillieren hinreichend viele Zentren in hinreichend vielen Arealen im Takt, dann erreicht das rhythmische Gesamtsignal das Bewußtsein und wird von den Belohnungssytemen mit einem guten Gefühl benotet und belohnt. Solche Großoszillationen im Cortex lassen sich auf der Schädelhaut → EEG als 40-80 Hertz Schwingung messen und mit fMRT lokalisieren.
Hierzu: Hirnforschung und Meditation: Ein Dialog.
Assoziatives Lernen
Schwingen Aktivitätszentren oft und lange synchron, dann ändert sich die Anatomie und es entstehen neue synaptische Verbindungen zwischen diesen Zentren. Sie synchronisieren sich in Zukunft leichter und schneller und es steigt die Tendenz zur gemeinsamen Aktivität.
| Vermutungen über den corticalen Algorithmus in einer Macrosäule: | |
| 1) | Die ankommenden Signale werden daraufhin untersucht, ob sie Ähnlichkeiten mit Prototypen haben. |
| 2) | Falls Ähnlichkeit mit einem Prototypen vorhanden, dann wird |
| 2a) an die nächst höhere i+1 Hierarchie- und Abstraktionsebene das Kennsignal des Prototypen und der Grad der Ähnlichkeit gemeldet und | |
| 2b) an die nächst tiefere i-1 Hierarchie- und Abstraktionsebene der Prototyp abgespielt wie eine Tonkonserve. | |
| 3) | Falls keine Ähnlichkeit vorhanden, wird |
| 3a) das ankommende Signal zur nächst höheren i+1 Ebene unverändert durchgereicht und | |
| 3b) die i-1 Ebene darüber informiert, dass man mit dem Signal nichts anfangen konnte. | |
| 4) | Die Kennsignale der Prototypen = Musternummern sind schlanker (enthalten weniger Redundanz und kein Rauschen) als das Eingangssignal. Das Erkennen eines Prototypen entlastet die nächst höhere Ebene i+1 vom Zwang, große Datenmengen annehmen zu müssen. |
| 5) | Signale werden also die Hierarchieebenen soweit hochgeschleust bis sie vollständig in Form von Prototypen-Codes codiert worden sind. |
| 6) | Signale, bei denen das durch alle Hierarchiestufen nicht gelingt, gehen in den Hippocampus, der für einige Zeit alles speichert, was neu- und fremdartig ist. Diese Inhalte tauchen in unserem Bewusstsein auf mit einer emotionalen Bewertung von neugierig, unbehaglich oder alarmierend. |
 |
Auch wenn die vorherigen Kapitel etwas anderes suggerieren, wissen wir über das Gehirn fast nichts. Jedenfalls ist unser Wissen verzweifelnd gering verglichen mit unserem Unwissen. |
Gedankenexperiment damit wir bescheiden bleiben:
Das Gehirn einer Biene hat ca. 100 Millionen Zellen. Damit kann die Biene bei Sonnenuntergang und Abendwind über einer blühenden Wiese fliegen ohne irgendwo anzustoßen, kann eine im Wind schwankende Pflanze auswählen und in einem einzigen Anflug auf der Blüte elegant landen und sich festhalten.
Leider kann niemand einen Autopiloten für einen Hubschrauber so programmieren, dass er auch nur ansatzweise das leistet, was dieses Insektengehirn leistet.
Noch auf Jahrzehnte wird diese Aufgabe unlösbar bleiben, weil unsere besten Computer einem Insektengehirn nicht das Wasser reichen können.
Vorsicht vor massenhaftem dummem Gerede in Presse und TV !
Dort behaupten gehirnlose Leute, sie hätten das Gehirn verstanden und sie würden ein Computer-Gehirn bauen.
| Erforschung des Cortex: | |
| Ramon y Cajal 1909, Barcelona | → Wikipedia |
| Vernon Mountcastle 1978, Baltimore | → Wikipedia |
| David Hubel & Torsten Wiesel 1981, Harvard | → Wikipedia |
| Erich Kandel 2000, New York | → Wikipedia |
| Jeff Hawkins 2004 | → www.onintelligence.org |
Literatur:
1) KANDEL, Eric: In Search of Memory, 2007 → www2.wwnorton.com/catalog/fall06/032937.htm
Deutsche Übersetzung: Auf der Suche nach dem Gedächtnis → www.randomhouse.de/..
2) HAWKINS, Jeff: On Intelligence, 2005 → search.barnesandnoble.com
Deutsche Übersetzung: Die Zukunft der Intelligenz, 2007 → www.rowohlt.de
| top of page: |